
For enquiries please contact us at kbrucksch@hotmail.com or kbrucksch@gmail.com.
Amber

Gladiator (?), Hukawng Valley, Myanmar, copyright by Alex Beigel.

Large pseudoscorpion in Myanmar amber

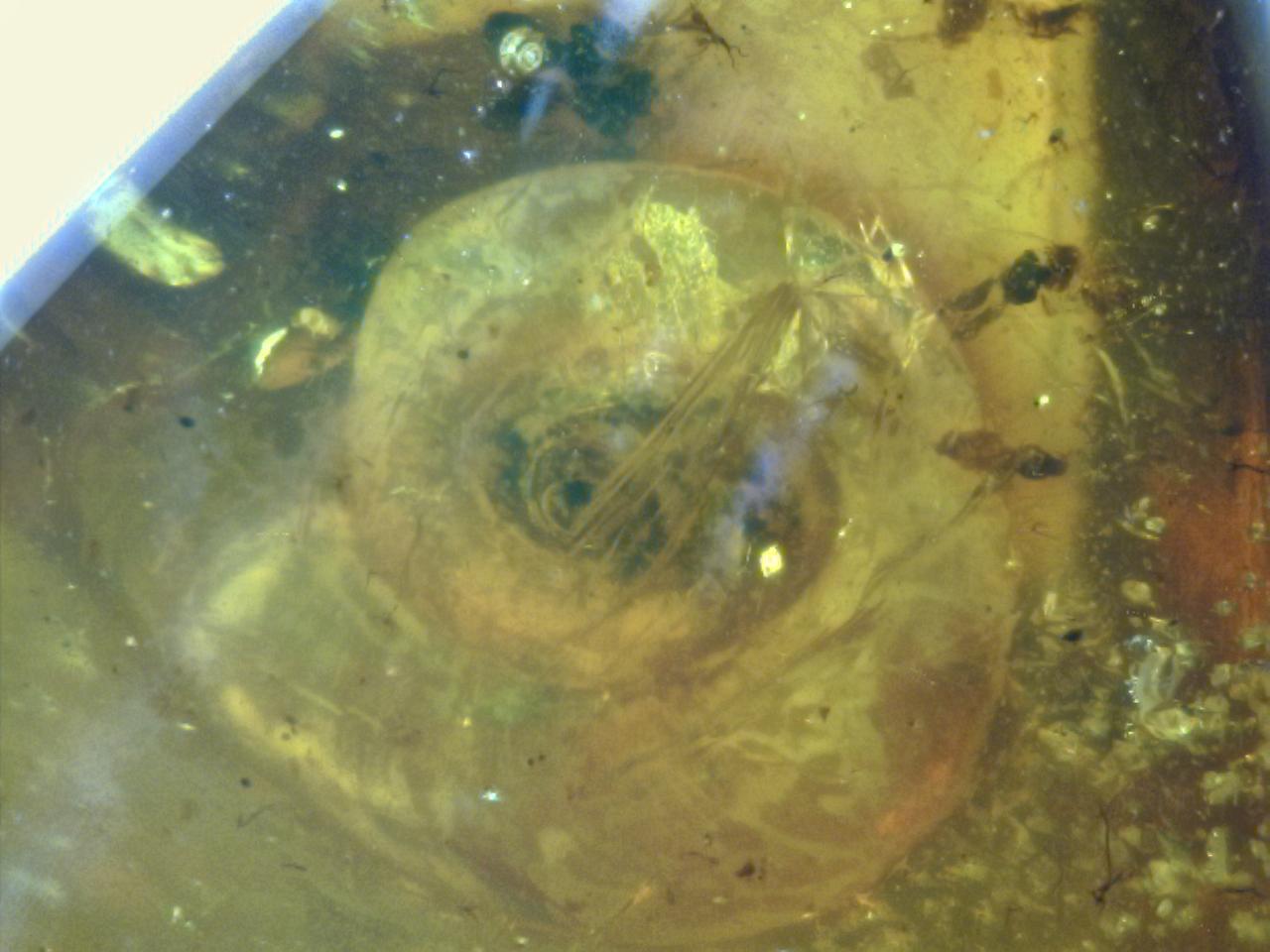
Chaerilobuthus bruckschi
A rare scorpion , Hukawng Valley, Myanmar.

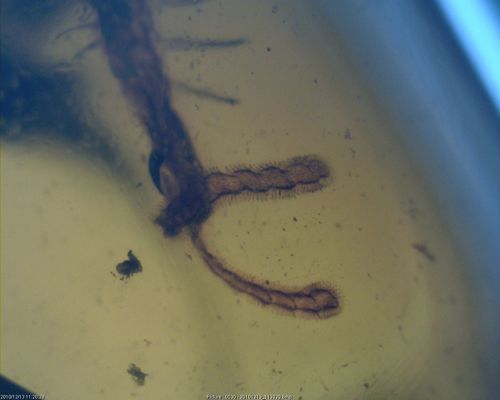
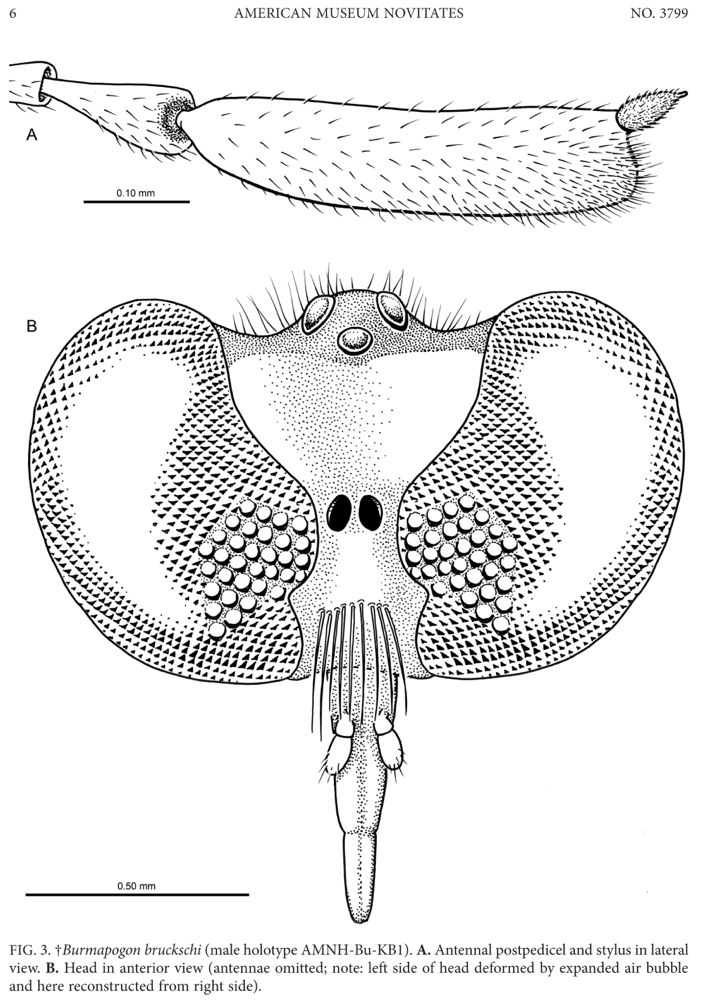
Burmapogon bruckschi
New species of robber fly, Hukawng Valley, Myanmar.
A snail shell, Hukawing Valley, Myanmar.
Amber (or resinite) is fossilised tree resin (not sap), which has been appreciated for its colour and natural beauty since Neolithic times. In the Greek mythology amber was believed to be tears of the Heliades, the daughters of the sun god.
When the volatile liquids evaporate the resin begins to polymerise. After 100s or 1000s of years it becomes ‘copal’ – also called ‘young amber’ – and after millions of years the resin becomes amber.
The presence of insects in amber was noticed by Pliny the Elder in his Naturalis Historia and led him to the (correct) theory that at some point, amber had to be in a liquid state to cover the bodies of insects.


Amber specimens
Amber from the Baltic Sea has been extensively traded since antiquity and in the main land, from where amber was traded 2000 years ago, the natives called it glaes (referring to its see-through similarity to glass).
The Baltic Lithuanian term for amber is Gintaras and LatvianDzintars. They and the Slavic jantar are thought to originate from Phoenician jainitar (sea-resin). However, while most Slavic languages, such as Russian and Czech, retain the old Slavic word, in the Polish language, despite still being correct, jantar is used very rarely (even considered archaic) and was replaced by the word bursztyn (burn stone) deriving from the German analogue (Bernstein).



Baltic sea amber inclusions.
Amber occurs in a range of different colours. As well as the usual yellow-orange-brown that is associated with the colour “amber”, amber itself can range from a whitish colour through a pale lemon yellow, to brown and almost black. Other more uncommon colours include red amber (sometimes known as “cherry amber”), green amber, and even blue amber, which is rare and highly sought after.
Much of the most highly-prized amber is transparent, in contrast to the very common cloudy amber and opaque amber. Opaque amber contains numerous minute bubbles. This kind of amber is known as “bony amber”.
Although all Dominican amber is fluorescent, the rarest Dominican amber is blue amber. It turns blue in natural sunlight and any other partially or wholly ultraviolet light source. In long-wave UV light it has a very strong reflection, almost white. Only about 100 kg is found per year, which makes it valuable and expensive.


Amber inclusions from Hukawng Valley, Myanmar.
The oldest amber recovered dates to the Upper Carboniferous period (320 million years ago). Its chemical composition makes it difficult to match the amber to its producers – it is most similar to the resins produced by flowering plants, which did not evolve until the Jurassic, around 180 million years ago. Amber becomes abundant soon afterwards, in the Early Cretaceous, 150 million years ago, when it is found in association with insects.
Commercially most important are the deposits of Baltic and Dominican amber.



Amber inclusions from various locations.
Baltic amber or succinite (historically documented as Prussian amber) is found as irregular nodules in marine glauconitic sand, known as blue earth,occurring in the Lower Oligocene strata of Samland in Prussia (Latin: Sambia), in historical sources also referred to as Glaesaria. After 1945 this territory around Königsberg was turned into Kaliningrad Oblast, Russia, where it is now systematically mined. It appears, however, to have been partly derived from yet earlier Tertiary deposits (Eocene); and it occurs also as a derivative mineral in later formations, such as the drift. Relics of an abundant flora occur as inclusions trapped within the amber while the resin was yet fresh, suggesting relations with the flora of Eastern Asia and the southern part of North America.


Amber inclusions from the Baltic sea and Mexico.
Amber can be found in many parts of the world ( Baltic Sea, Borneo, Canada, China, Dominican Republic, France, Italy, Japan, Lebanon, Mexico, Myanmar, Portugal, Romania, Russia, Spain). The best samples with inclusions though come from the Baltic region, Dominican Republic and Myanmar.
More interesting reading on:
I have many interesting inclusions in mainly Myanmar amber for sale. For purchase please contact me at kbrucksch@hotmail.com. My friend, Alex, has taken some excellent photos of my insect inclusions. Below are a few examples.

Larva, Hukawng Valley, Myanmar.

Assassin spider ‘Archaeidae’, Hukawng Valley, Myanmar.

Spider, Hukawng Valley, Myanmar.



2 baltic amber and a Colombian copal inclusions


















Myanmar amber inclusions

How beautiful I am.
Chaerilobuthus bruckschi
Clarification of the familial status of the genus Palaeoburmesebuthus Lourenço, 2002 from Cretaceous Burmese amber (Scorpiones: Archaeobuthidae: Palaeoburmesebuthinae)
Wilson R. Lourenço Muséum national d’Histoire naturelle, Département Systématique et Evolution, UMR7205, CP 053, 57 rue Cuvier, 75005 Paris, France: e-mail: arachne@mnhn.fr
Abstract: The study of five new scorpion specimens from the Cretaceous amber of Myanmar (Burma) lead to the description of three new species and to the clarification of the status of the genus Palaeoburmesebuthus Lourenço, the first scorpion described from Burmese amber. To present, eight species and eleven fossil scorpions have been described from Burmese amber. This attests to a considerable degree of diversity in the Burmese amber-producing forests. Key-words: scorpion, fossil, new subfamily, new species, Cretaceous, Burmese amber, Myanmar.
Introduction
After the conclusion of my previous article (this volume), I had the opportunity to examine five new Burmese amber pieces containing scorpions. Their study leads to the description of three more new species belonging to previously described genera. More significantly, however, are the new taxonomic elements furnish by one of the specimens allowing the clarification of the familial status of the genus Palaeoburmesebuthus Lourenço. Palaeoburmesebuthus grimaldii Lourenço, 2002 was the first scorpion described from Burmese amber (Lourenço, 2002). However, in account of the incompleteness of the specimen used in the description, P. grimaldii was placed as incertae familiae. A redescription was also published for Palaeoburmesebuthus grimaldii, providing some new insights to the taxa (Santiago-Blay et al., 2004). However, the familial status of this genus remained unclear.
The new species of Palaeoburmesebuthus described here shows taxonomic characteristics allying it to the extant family Buthidae, but also to the Cretaceous family Archaeobuthidae Lourenço, 2001, described from Lebanon amber (Lourenço, 2001). For this reason, it is placed in the family Archaeobuthidae, but attributed to a new subfamily, which is described here.
Chaerilobuthus bruckschi sp. n (Figs. 15-23)
Holotype is a juvenile specimen, also slighly larger than C. complexus, the first species described for this genus (Lourenço & Beigel, 2011) with 8.75 mm in total length. Sex cannot be determined but the morphology of the body and appendages may suggest a male. The block of amber is very clear, pale yellow and measured 22 x 14 x 7 mm. However, most characters are not observable because the specimen suffered with the dissection process within the resin, is twisted and located in a zone of fracture. The general morphology of the scorpion clearly associates it to the genus Chaerilobuthus. Type locality and horizon: Myanmar (Burma), Kachin; precise locality unknown; Lower Cretaceous. Patronym: The specific name honours Mr Klaus-Peter Brucksch, Kuranda, Australia, who arranged facilities for the study of the holotype. Depository: The type specimen is deposited in the personal collection of Mr Klaus-Peter Brucksch, Kuranda, Australia.
Diagnosis: General morphology shows similarities with both buthid and chaerilid scorpions and the specimen can be associated to the genus Chaerilobuthus. The following combination of features can be used to diagnose the new species: Carapace smooth, apparently with inconspicuous median eyes; a few trichobothria observed which can relate it to the pattern previously observed for Chaerilobuthus; at least 2 external trichobothria on the patella; 3-4 trichobothria on the dorso-external aspect of chela hand, two trichobothria at the base of fixed finger and two trichobothria on fixed finger. Fixed and movable fingers of pedipalp chela each with 6/7 series of small rounded granules separated by conspicuous accessory granules. Telson with a bulbous vesicle: aculeus moderately long and curved; base of aculeus not enlarged. Tibial spurs absent from legs III and IV.
Description: Coloration: the scorpion is reddish-brown to dark brown, probably due to the process of dissection suffered by the specimen. The ventral aspect of the specimen cannot be clearly observed since it is located in the zone of fracture of the piece.
Morphology. Carapace apparently smooth, not granular; anterior margin almost straight. Carinae and furrows absent. Median ocular tubercle more or less distinct; median eyes, if present, are very small. Lateral eyes not observable. Sternum not observable. Mesosomal tergites not granular and acarinate. Ventral aspect not well observable. Metasomal segments I to IV with 10-8-8-8 carinae; segment V with five carinae; all carinae conspicuous; dorsal carinae of segments I-IV with a series of spinoid granules; dorsal aspect of segments I-V strongly depressed; setation on segments I to V moderately to strongly marked. Telson with a bulbous vesicle, not flattened dorso-ventrally; weakly granular to smooth; aculeus moderately long and curved. Cheliceral dentition not visible.
Pedipalp femur probably pentacarinate; patella with 5-6 carinae. Chela with moderately marked carinae; all faces almost smooth. Fixed and movable fingers with 6-7 rows of small, rounded granules, separated by strong spinoid accessory granules; extremity of fingers without any stronger spinoid granule. Trichobothriotaxy: trichobothrial pattern only partially observed (see diagnosis). Leg: tibial spurs absent.
Morphometric values (in mm) of the juvenile holotype of Chaerilobuthus
bruckschi sp. n.
Total length 8.75 (including telson). Carapace: length 1.03, posterior width -. Mesosoma length 3.11. Metasomal segments length/width: I 0.40/0.33; II 0.47/0.27; III 0.53/0.27; IV 0.67/0.27; V 1.20/0.25. Telson length/width/height: 1.34/0.42/0.47. Pedipalp: patella length 1.46, width 0.52; chela length 1.87, height 0.34; movable finger length 0.93.
Burmapogon bruckschi
To read the whole publication of the American Museum click here .



Leviunguis bruckschi



